
Sur une photographie aujourd’hui célèbre, un homme au visage émacié, vêtu d’une toge universitaire et d’une curieuse écharpe à pompons, pose fièrement. Sa main non gantée repose en un geste propriétaire sur le squelette d’un animal sans ailes haut de trois mètres. Cet homme à l’air peu commode, c’est Richard Owen, un paléontologue britannique du XIXème siècle, célèbre, entre autres choses, pour avoir inventé le mot « dinosaure ». Il pose ici aux côtés du squelette d’un moa, un oiseau endémique à la Nouvelle-Zélande, éteint avant l’arrivée des Occidentaux.
Les moa1 sont, avec les « oiseaux éléphants » de Madagascar les représentants les mieux connus de ce qu’on appelle couramment les « oiseaux géants ». Cette expression rassemble des espèces de diverses époques et régions du monde et qui n’ont parfois que des liens éloignés du point de vue de l’évolution. Certaines comme les émeus vous sont sans doute déjà familières mais bien d’autres ont jadis foulé les sols du Pacifique et se sont éteintes depuis. Casoar vous emmène aujourd’hui à la découverte de ces grands oiseaux disparus.

Richard Owen posant aux côtés d’un squelette de Dinornis robustus, 1879
Pour bien commencer notre voyage dans le temps, il nous faut nous armer des outils des sciences naturelles, en particulier de la classification phylogénétique. Derrière cette expression se cache une certaine méthode d’organisation, de classement, des êtres vivants peuplant ou ayant peuplé la terre. La classification phylogénétique, parfois aussi appelée cladistique, classe les êtres vivants en fonction de leur degré de parenté dans l’évolution. Elle s’organise ainsi en arbres phylogénétiques qui fonctionnent un peu comme des arbres généalogiques à l’échelle des espèces. Pour déterminer si deux espèces sont apparentées ou non, on compare leurs différentes caractéristiques morphologiques. En effet, on considère que des caractéristiques morphologiques communes peuvent avoir été héritées d’un ancêtre commun. En se basant sur ces comparaisons, on va pouvoir ranger les différentes espèces dans des boîtes appelées taxons. La première boîte, la plus petite, c’est « l’espèce ». Les espèces qui présentent un certains nombre de points communs, et donc qu’on suppose avoir un ancêtre commun, forment un « genre » et les genres qui ont un certain nombre de points communs entre eux forment une « famille », les familles forment des « ordres » qui forment eux-même des « classes » et ainsi de suite. À chaque fois la boîte est de plus en plus grande et l’ancêtre commun est de plus en plus éloigné2.
Les résultats de cette méthode peuvent parfois être démentis par l’étude de la génétique des êtres vivants. En effet, il arrive que deux espèces présentent une même caractéristique morphologique mais qu’elles ne possèdent pas d’ancêtre commun : c’est ce qu’on appelle une convergence évolutive. Par ailleurs, de nouvelles études du matériel disponible peuvent toujours remettre en cause les classifications établies. Néanmoins, cette méthode est très utile en paléontologie, où les chercheurs ne peuvent pas toujours avoir recours aux analyses génétiques3.
Ce qu’il est important de comprendre ici c’est qu’il n’existe pas une classification qui serait définitive ou détentrice d’une vérité ultime. Toutes les classifications du vivant, anciennes ou contemporaines, sont le fruit du regard que l’être humain porte sur le monde qui l’entoure et reflètent un certain état culturel et un certain état des sciences. Quoiqu’on ait tendance à penser, les sciences sociales ne sont pas les seules à être influencées par la culture des groupes humains qui les produisent, les sciences naturelles elles aussi évoluent au gré des changements culturels. Par exemple, au XVIIIème siècle il était fréquent que les ossements de dinosaures qui pouvaient parfois être retrouvés par hasard soient interprétés comme ayant appartenu à des géants antédiluviens, décrits dans la Bible, venant ainsi confirmer la version chrétienne de l’origine du monde et des espèces4.
Revenons à nos oiseaux et essayons de rendre tout cela un peu plus concret. Quand Owen invente le terme « dinosaure », qui signifie « lézard terriblement grand », il pense que les fossiles qu’il étudie ont un lien de parenté évolutif avec les reptiles actuels. En fait, les dinosaures sont d’un point de vue évolutif bien plus proches des oiseaux que des crocodiles ou des lézards. Selon la classification phylogénétique, les oiseaux sont des dinosaures. Si on repense à la photo d’Owen posant aux côté du moa le retournement à quelque chose d’ironique : le père des dinosaures finalement rattrapé par son sujet d’étude.
Ces points généraux posés, nous pouvons embarquer pour le Pacifique, il y a quelques centaines d’années. Imaginez que vous êtes dans une forêt de Nouvelle-Zélande. Vous vous promenez bien gentiment quand soudain, au détour d’un tronc d’arbre apparaît un drôle d’oiseau. La créature fait environ deux mètres de haut, pèse un bon quart de tonne et est couverte des pieds à la tête de plumes brunes. Elle se tient sur ses deux pattes arrières, ses deux seules pattes, car vous remarquez rapidement qu’elle est totalement dépourvue d’ailes. Son cou n’est pas vertical mais plutôt horizontal et forme un creux important à sa base. Vous êtes face à un Dinornis robustus, l’une des neuf espèces de moa aujourd’hui répertoriées5.
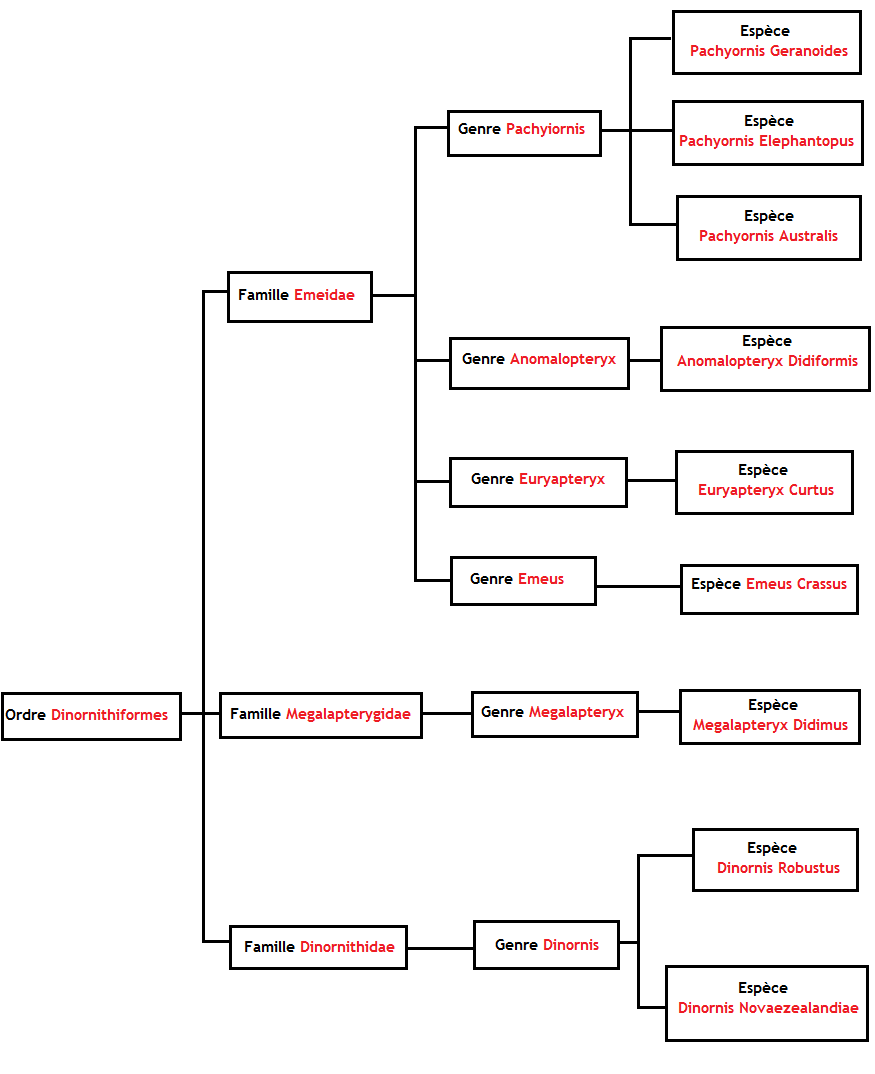
Arbre phylogénétique des moa (la classification retenue est celle proposée par SCOFIELD, R & WORTHY, T., 2012. « Twenty-first century advances in knowledge of the biology of moa (Aves : Dinornithiformes) : a new morphological analysis and moa diagnoses revised » in New Zealand Journal of Zoology, 39:2. Wellington, Royal Society of New Zealand. p. 87-153.)
Ces neuf espèces se répartissent en six genres eux mêmes répartis en trois familles qui forment l’ordre des dinornithiformes. Moa est un terme d’origine māori qui a été adopté au XIXème siècle par les Occidentaux pour désigner couramment l’ensemble de ces neuf espèces. La classification phylogénétique a depuis permis aux paléontologues d’établir que les moa étaient proches des kiwi et des anciens oiseaux éléphants de Madagascar mais également des casoar. Ils sont aussi des parents, plus éloignés, des autruches.
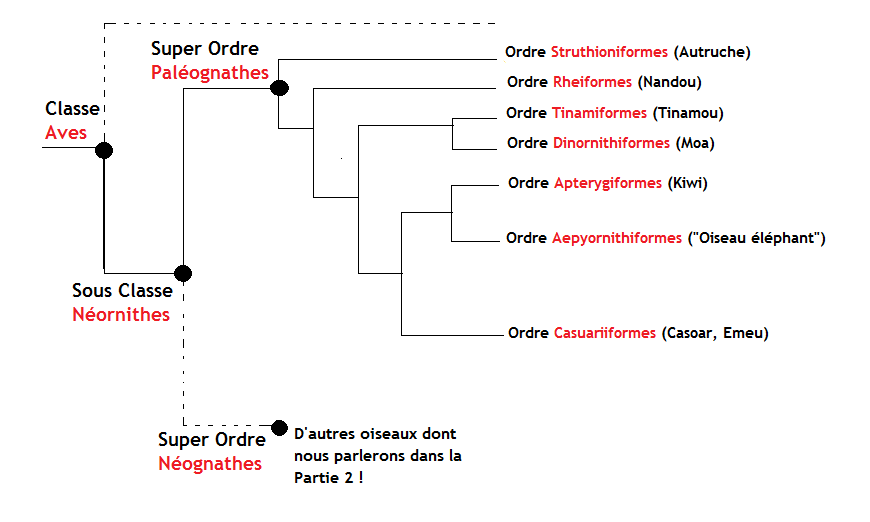
Arbre phylogénétique simplifié des moa montrant leurs liens avec les autres espèces de grands oiseaux : les taxons (ou clades) sont indiqués en noir, chaque embranchement indique une évolution à partir d’un ancêtre commun. Par exemple on peut voir ici que les moa partagent un ancêtre commun avec les tinamou, des oiseaux d’Amérique du sud. Tinamou et moa partagent également un ancêtre commun plus éloigné avec les casoars, les émeus, les oiseaux éléphant et les kiwi. (cet arbre s’inspire très largement de celui fourni par ANGST, D & BUFFETAUT, E., 2018. Paléobiologie des oiseaux géants terrestres. London, ISTE Editions.)
C’est au XIXème siècle que les scientifiques occidentaux commencent à s’intéresser aux ossements fossiles de grande taille découverts lors de travaux d’aménagement tels que le drainage de certains marais. Nombre de ces fossiles sont envoyés en Angleterre et se retrouvent dans le bureau de Richard Owen, véritable pape de la paléontologie britannique de l’époque. En paléontologie, le premier à publier la description d’une nouvelle espèce à partir de ses restes fossilisés gagne le droit de la nommer et c’est donc à Owen que l’on doit la plupart des noms latins des espèces de moa.
Les esprits s’emballent cependant quelque peu, les fossiles s’accumulent et les scientifiques décrivent pas moins de vingt-neuf espèces de moa différentes ! Il peut en effet arriver aux paléontologues de penser qu’ils ont entre les mains un fossile appartenant à une nouvelle espèce alors qu’il s’agit en fait d’un membre d’une espèce déjà connue. La paléontologie travaille souvent à partir de squelettes incomplets et d’ossements fragmentaires, il est donc aisée de faire des erreurs. Un cas de figure fréquent est par exemple la description de fossiles comme une nouvelle espèce alors qu’ils appartiennent en fait à un jeune individu d’une espèce déjà référencée et apparaissent donc au premier abord comme différents.
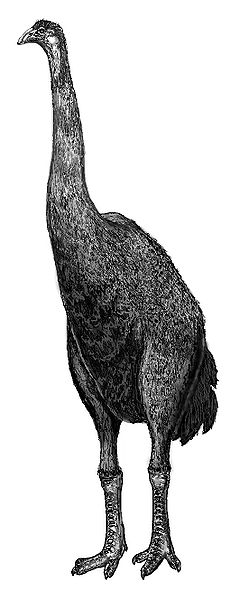
Représentation ancienne d’un moa : le cou est présenté à la verticale
Il faut également savoir qu’au XIXème siècle et au début du XXème siècle il existe une sorte de course entre les paléontologues : c’est à qui décrira le plus de nouvelles espèces. Or, comme on l’a expliqué plus haut, premier arrivé, premier servi ! Si un paléontologue décrit une espèce déjà décrite et publiée par un collègue, même un seul jour avant, on retiendra le collègue comme découvreur de l’espèce en question. Cet état de fait a parfois pu amener les scientifiques de l’époque à décrire des espèces à partir de fossiles extrêmement incomplets afin de couper l’herbe sous le pied de leurs confrères. L’exemple le plus célèbre est la fameuse « guerre des os » que se sont menés les paléontologues Edward Cope et Othniel Marsh aux Etats-Unis à la fin du XIXème siècle et qui avait pour objet des ossements de dinosaures. Cet épisode, quoique fort divertissant, est cependant un peu trop éloigné de notre propos pour que nous nous y attardions.6 Aujourd’hui, l’ADN est particulièrement utile pour débrouiller ces débats et c’est lui qui a permis de réduire le nombre d’espèces de moa à seulement neuf.
Les études de la seconde moitié du XXème siècle et du début du XXIème ont également permis de rectifier d’autres erreurs du XIXème siècle. Vous souvenez-vous de la photo d’Owen posant aux côté de son squelette de moa au cou fièrement dressé? On sait aujourd’hui que les moa ne portaient pas leur cou à la verticale mais sans doute plutôt à l’horizontal comme on l’a dit plus haut. Au XIXème siècle cependant, on pensait que les moa avaient un cou beaucoup plus droit ce qui a amené les scientifiques à surestimer leur taille. On parle ainsi parfois de moa hauts de pas moins de 3 à 4m. Les paléontologues estiment désormais que cette taille, qui a fait la célébrité des moa, est largement exagérée. Premièrement, toutes les espèces ne faisaient pas la même taille et il existait donc de petits moa. L’espèce Anomalopteryx didiformis ne dépassait par exemple vraisemblablement pas les 90 cm de hauteur. La plus grande espèce était Dinornis robustus dont les femelles pouvaient mesurer jusqu’à 2m de haut et peser plus de 200kg. Petit détail intéressant, les mâles de cette espèce étaient jusqu’à quatre fois plus légers et beaucoup plus petits. C’est sans doute eux qui couvaient les œufs, les femelles étant trop lourdes.

Squelette de moa de l’espèce Megalapteryx didinus présenté à l’Otago Museum de Dunedin : vous remarquerez le cou orienté à l’horizontal
Contrairement à ce qu’on peut voir sur de nombreuses représentations anciennes, les moa vivaient dans un milieu boisé et non pas dans les vastes plaines de la Nouvelle-Zélande contemporaine. Ces plaines sont en effet le fruit des activités humaines à la fois récentes et anciennes qui ont considérablement modifié le paysage. Les moa vivaient donc plutôt dans des forêts et n’étaient vraisemblablement pas des coureurs rapides, contrairement encore une fois à ce qu’on peut voir représenté sur certaines illustrations. La forme de leurs pattes suggèrent en effet qu’ils se déplaçaient plutôt lentement.
On ne sait évidemment pas grand chose de plus sur le mode de vie de ces oiseaux mis à part qu’ils étaient herbivores et étaient sans doute la proie favorite de l’aigle de Haast. Cet aigle endémique à la Nouvelle-Zélande pouvant mesurer jusqu’à 3m d’envergure pour les femelles et peser une quinzaine de kilos n’hésitait pas à s’attaquer aux plus grosses espèces de moa. Il s’agit du plus gros rapport entre le poids d’un prédateur et celui de sa proie chez les vertébrés terrestres. Comme pour le moa, on ne connaît cet oiseau éteint que grâce à des fossiles découverts sur l’île du sud de Nouvelle-Zélande. Il doit son nom à Julius von Haast, un géologue allemand installé en Nouvelle-Zélande et le fondateur du Canterbury Museum de Christchurch.7

Moa attaqué par un aigle de Haast : reconstitution proposée par le Te Papa Tongarewa de Wellington
Il reste un point que nous n’avons pas abordé, c’est celui de la chronologie. On estime actuellement que l’histoire évolutive des moa aurait débuté durant le Miocène, une ère géologique qui débute il y a environ 23,5 millions d’années pour se terminer il y a 5,3 millions d’années8. Leur extinction, à la fois sa cause et sa chronologie, a quant à elle fait l’objet de très nombreux débats et de théories, pour certaines fort alambiquées. Cette interrogation est sans doute l’une des sources de la fascination pour les moa et de la très abondante littérature qui existe à leur sujet. Depuis le XIXème siècle les scientifiques ont ainsi eu tout le temps de répondre à cette grande question : pourquoi et quand les moa ont-ils disparus?
Pour connaître la réponse et en découvrir plus sur les grands oiseaux disparus du Pacifique rendez-vous la semaine prochaine sur Casoar !
Alice Bernadac
Image à la une : Moa attaqués par un aigle de Haast, illustration réalisée par John Megahan
1 Moa étant un terme d’origine māori il ne sera jamais accordé dans cet article.
2 Les systématiciens qui passeront par ici devront me pardonner pour cette explication succincte qui ne rend justice ni à l’histoire de la cladistique, ni aux débats qui peuvent l’entourer et que je maîtrise très insuffisamment. Il s’agit ici d’un article de vulgarisation qui vise à expliquer les principes généraux de cette méthode et à donner envie d’aller lire des personnes plus qualifiées que moi sur le sujet.
3 Pour plus d’informations sur la cladistique et l’évolution des espèces en général on pourra consulter LECOINTRE, G. & LE GUYADER, H., 2016. Classification phylogénétique du vivant Tome 1 (4e édition). Paris, Belin. ainsi que LECOINTRE, G (dir.) et al., 2009. Guide critique de l’évolution. Paris, Belin.
4 Pour un ouvrage abordant de façon très accessible l’histoire de la paléontologie voir LE LOEUFF, J. 2016. T-Rex Superstar. Paris, Belin.
5 Les informations qui vont suivre sur les caractéristiques physiques des moa proviennent de ANGST, D & BUFFETAUT, E., 2018. Paléobiologie des oiseaux géants terrestres. London, ISTE Editions.
6 Pour plus d’informations sur cet épisode voir ALLAIN, R., 2015. Histoire des dinosaures. Paris, Flammarion.
7 Sur l’aigle de Haast voir HOLDAWAY, R. & WORTHY, T., 2002. The lost world of the moa. Bloomington, Indiana University Press. ainsi que ANGST, D & BUFFETAUT, E., 2018. Paléobiologie des oiseaux géants terrestres. London, ISTE Editions.
8 Suivent ensuite le Pliocène (5,3 Ma- 1,8 Ma), le Pléistocène (1,8 Ma – 0,01 Ma) et l’Holocène (0,01 Ma – ?) (Certains scientifiques considèrent aujourd’hui qu’une nouvelle ère géologique, l’Anthropocène, a débuté au XIXe siècle au moment de la révolution industrielle mais nous n’entrerons pas ici dans les débats qui entourent cette notion)
Bibliographie :
- ANDERSON, A., 1989. Prodigious birds : moas and moas hunting in prehistoric New Zealand. Cambridge, Cambridge University Press.
- ANGST, D & BUFFETAUT, E., 2018. Paléobiologie des oiseaux géants terrestres. London, ISTE Editions.
- HOLDAWAY, R. & WORTHY, T., 2002. The lost world of the moa : prehistoric life of New Zealand. Bloomington, Indiana University Press.
- SCOFIELD, R & WORTHY, T., 2012. « Twenty-first century advances in knowledge of the biology of moa (Aves : Dinornithiformes) : a new morphological analysis and moa diagnoses revised » in New Zealand Journal of Zoology, 39:2. Wellington, Royal Society of New Zealand. p. 87-153.
3 Comments